


تحقیق بر روی RNA منجر به کشف های زیست شناختی مهم و جوایز نوبل متعدد شده است. نوکلئیک اسید ها در سال ۱۸۶۸ توسط Friedrich Miescher کشف شد؛ کسی که ماده نوکلئین را به دلیل این که در هسته پیدا شد، به این عنوان نامگذاری کرد. بعداً کشف شد که سلول های پروکاریوتی که فاقد هسته هستند، نیز حاوی اسید های نوکلئیک هستند. نقش RNA در سنتز پروتئین همواره از سال ۱۹۳۹ مورد توجه بود. Severo Ochoa جایزه نوبل را(مشترکاً با Arthur Kornberg) در سال ۱۹۵9، بعد از کشف آنزیمی که RNA را در آزمایشگاه سنتز می کرد، برنده شد. بعد ها نشان داده شد که آنزیم کشف شده توسط Severo Ochoa (polynucleotide phosphorylase)، مسئول تجزیه RNA باشد نه آن که RNA را سنتز کند. در سال ۱۹۶5، توالی ۷۷ نوکلئوتیدی از یک tRNA مخمر توسط Robert W. Holley (برنده جایزه نوبل ۱۹۶۸در داروسازی، مشترکاً با Har Gobind Khorana و Marshall Nirenberg) کشف شد. در سال۱۹۶۷، Carl Woese فرضیه داد که RNA ممکن است کاتالایتیک باشد و ابتدایی ترین اشکال حیات (مولکول های خودرونوشتبردار) توانستند برای حمل اطلاعات ژنتیکی شان و هم برای کاتالیز واکنش های بیوشیمیایی شان به RNA تکیه کنند.
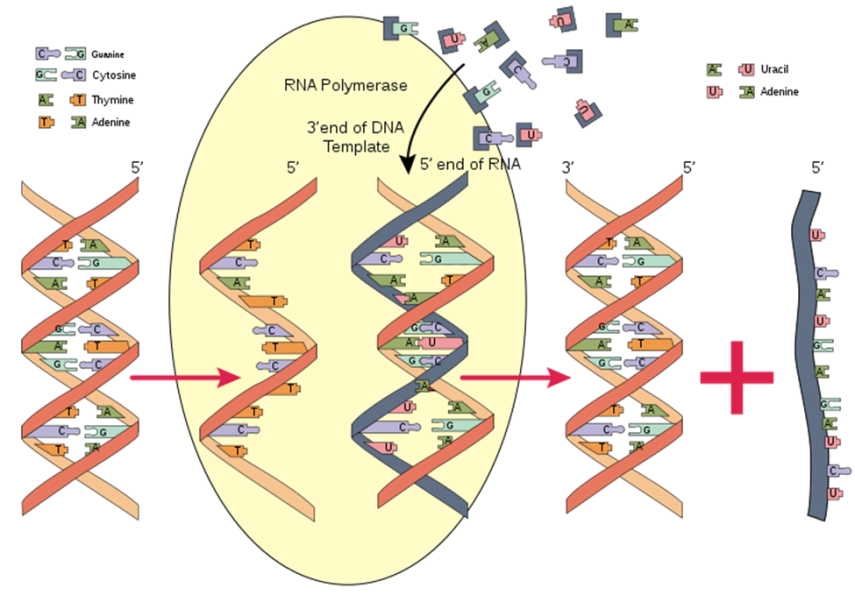
مقایسه ساختمان RNA با DNA
RNA و DNA هر دو نوکلئیک اسید هستند، اما در سه چیز تفاوت دارند:
نخست این که، برخلاف DNA که دو رشته ای است، RNA یک مولکول تک رشته ای است و زنجیره بسیار کوتاه تری از نوکلئوتید ها را دارد.
دوم این که، در حالی که DNA دارای قند دئوکسی ریبوز می باشد، RNA دارای ریبوز است. این گروه های هیدروکسیل، پایداری RNA را کمتر از پایداری DNA می کنند، زیرا داشتن گروه هیدروکسیل، ریبوز را برای واکنش هیدرولیز آماده تر می سازد.
سوم این که برخلاف DNA در RNA، باز تکمیل کننده آدنین، تیمین نیست، بلکه یوراسیل می باشد که شکل متیله نشده ای از تیمین می باشد.
ساختار بیشتر RNA های کارا از دیدگاه زیستی که شامل RNA کوچک هسته ای، rRNA، tRNA، mRNA و دیگر RNA های بی رمز، گهگاه دارای چیدمان های خود تکمیل کننده ای هستند که به بخش هایی از RNA این اجازه را می دهند که با خودش جفت شده و تا بخورد و مارپیچ های دوتایی را پدید آورند(همانند DNA). برخلاف DNA، دارای مارپیچ های دوتایی دراز نیستند، اما در جای جای آن ها گروه هایی از مارپیچ های کوتاه دیده می شود.
گونه های RNA
mRNA، که داده ها را از DNA به ریبوزوم، جایگاه ساخت پروتئین و ترجمه در یاخته، می برد. چیدمان RNA پیام رسان، چیدمان اسید آمینه های پروتئینی که ساخته می شود را تعیین می کند.
بسیاری از RNA ها در ترجمه به پروتئین تبدیل نمی شوند. به این RNA ها، بی رمز می گویند که برجسته ترین نمونه RNA بی رمز، tRNA ها و rRNA هستند که هر دوی آن ها در فرآیند ترجمه دارای کارکرد می باشند.
rRNA ها می توانند واکنش های شیمیایی، مانند برش و اتصال دیگر مولکول های RNA، را انجام دهند و تشکیل پیوند پپتیدی را در ریبوزوم ها آسان کنند.
در فرآیند ترجمه، mRNA، داده های مورد نیاز در چیدمان اسید های آمینه یک پروتئین را به ریبوزوم ها می برند. پروتئین ها به گونه ای کد می شوند که هر سه نوکلئوتید(یک کدون) مطابق با یک اسید آمینه است. در سلول های یوکاریوتی، همین که mRNA پیش ساز (pre mRNA) از DNA رونویسی شد، به mRNA بالغ پردازش می شود. در این فرآیند، اینترون های (بخش های کد نشونده pre mRNA) آن جدا می شوند. mRNA ها سپس از هسته به سیتوپلاسم صادر می شوند، جایی که به ریبوزوم متصل می شوند و به کمک tRNA به شکل پروتئین متناظرش، ترجمه می شود.
در سلول های پروکاریوتی که هسته و اجزای سیتوپلاسمی ندارند، mRNA می تواند به ریبوزوم ها متصل شود، در حالی که از DNA رونویسی می شود. بعد از مقدار معینی گذشت زمان، mRNA به نوکلئوتید های مولفه خودش با یاری ریبونوکلئاز ها تجزیه می شود. RNA ناقل(tRNA)، یک زنجیره RNA کوچک با حدود ۸۰ نوکلئوتید است که یک اسید آمینه خاص را به زنجیره پلی پپتیدی در حال رشد در محل ریبوزومی سنتز پروتئین در طی ترجمه، منتقل می کند.
rRNA، جزء کاتالیتیک ریبوزوم ها می باشند. ریبوزوم های یوکاریوتی حاوی ۴ مولکول rRNA مختلف می باشند: rRNA های 18S، 28S، 5.8S، 5S. سه تا از مولکول های rRNA در هسته سنتز می شوند و rRNA باقی مانده در جای دیگر سنتز می شود. ریبوزوم به mRNA متصل می شود و سنتز پروتئین را اجرا می کند. چندین ریبوزوم ممکن است به یک mRNA منفرد در هر زمانی متصل شوند. rRNA بینهایت فراوان است و ۸۰٪ از (mg/ml ۱۰) RNA یافت شده در سیتوپلاسم یک نمونه یوکاریوتی را تشکیل می دهد.
RNA پیامبر-ناقل (tmRNA)، در بسیاری از باکتری ها و پلاستید ها یافت می شود. tmRNA، پروتئین های کد شده توسط RNA پیامبر(mRNA) را نشان دار می کند که فاقد کدون های توقف برای تجزیه هستند و ریبوزوم را از ماندن باز می دارد.
RNA های تنظیمی، چندین نوع از RNA می توانند بیان ژن را با مکمل یکدیگر بودن در یک بخش mRNA یا یک DNA ژن، کاهش دهند.
RNA های کوچک(miRNA)، با ۲۱ تا ۲۲ نوکلئوتید در یوکاریوت ها یافت می شوند و از طریق مداخلهRNA (RNAi) عمل می کنند. به این صورت که یک کمپلکس مؤثر از miRNA و آنزیم ها می توانند mRNA را به miRNA که مکمل است بشکند، mRNA را از ترجمه شدن ممانعت کند یا تجزیه آن را تسریع کند. در حالی که RNA های مداخله ای کوچک(siRNA; 20-25 nt)، غالباً توسط تجزیه RNA ویروسی تولید می شوند، منابع درون زادی نیز از siRNAها وجود دارد. siRNAها از طریق RNA مداخله گر در یک روش مشابه با miRNA عمل می کنند. بعضی از siRNA ها و miRNA ها می توانند ژن هایی را که آن ها نشان دار می کنند، متیله کنند که به این وسیله، رونویسی این ژن ها را کاهش یا افزایش می دهند.
جانوران، RNA های بر هم کنش دهندهPiwi (piRNA; 29-30 nt) دارند که درسلول های جنینی فعال هستند و تصور می شود که دفاعی در برابر ترانسپوزون ها(transposons) باشند و نقشی در گامتوژنز ایفا کنند.
بسیاری از پروکاریوت ها، RNA های CRISPR(یک سیستم تنظیمی مشابه باRNAi) دارند.
RNA های آنتی سنس، بسیار گسترده و وسیع هستند، اکثراً یک ژن را فروتنظیم می کنند، اما تعدادی نیز فعال کننده رونویسی هستند. یک روشی که RNA آنتی سنس می تواند عمل کند، از طریق اتصال به یک mRNA و تشکیل یک RNA دورشته ای می باشد که به طور آنزیمی تجزیه می شود.
RNA های غیر کدکننده طویل بسیاری وجود دارد که ژن ها را در یوکاریوت ها تنظیم می کنند، یکی از این RNA ها، xist است که یک کروموزوم x را در پستانداران ماده می پوشاند و آن را غیرفعال می کند.
یک mRNA ممکن است حاوی اجزای تنظیمی خود به خودی باشد، از جمله riboswitche ها، در ناحیه ترجمه نشونده ‘۵ یا ناحیه ترجمه نشونده ‘۳. این عناصر تنظیمی sis، فعالیت آن mRNA را تنظیم می کنند. ناحیه های ترجمه نشونده همچنین ممکن است حاوی عناصری باشند که دیگر ژن را تنظیم می کنند.



